Chapter 12 Biogeochemistry
12.1 Life on Earth
All oxygen on Earth arises from microbial photosynthesis. It is because of oxygen in the environment that Earth is habitable for humans and most other organisms that we know.
Microbes also drive the biogeochemical cycles that are needed for life: this is because biogeochemical cycles also allows for essential elements (e.g., carbon, hydrogen, oxygen, nitrogen, sulfur, and phosphorous).
The above cycles are a series of redox reactions that bacteria and archaea use for energy (i.e., e- acceptors and donors) to make organic carbon. The elements recycled also make up the biomolecules that make life (e.g., DNA, RNA, proteins, etc).
12.1.1 Hopanoids
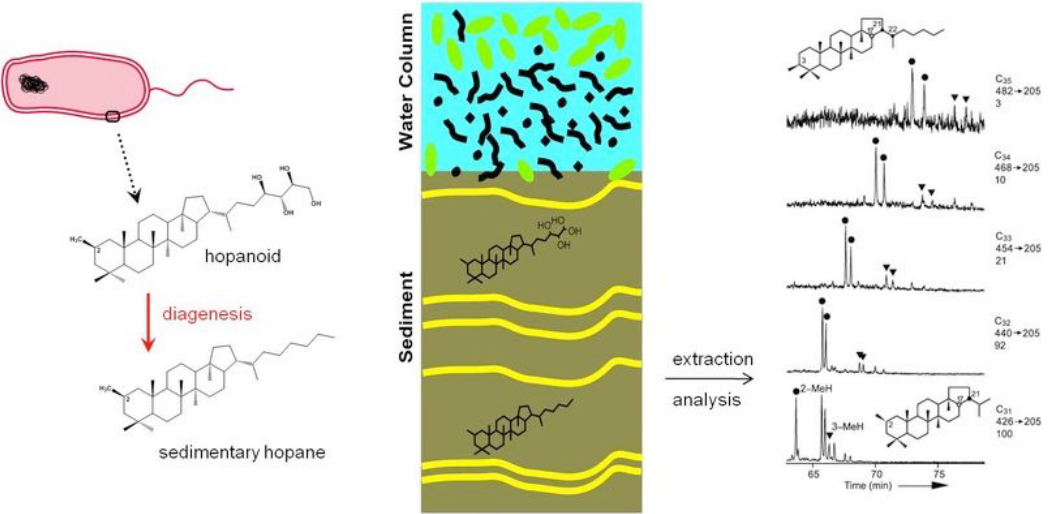
Figure 12.1: Hopanoid Analysis
These are molecules that provide evidence of photosynthesis prior to the oxygenation of the atmosphere.
Neverless, hopanoids are found in cell membranes and thought to play a role in membrane fluidity, stress, and protein localization.
Just like DNA, RNA, and proteins, hopanoids can be used as biomarkers. Hopanoids are also much more stable than DNA, RNA, and proteins and are hence used as biomarkers in fossils. 2-methylhopanoids are used as biomarkers for cyanobacteria and oxygenic photosynthesis, but it is crucial to note that hopanoids do not contain genetic information and are therefore harder to identify!
12.2 Phototrophy
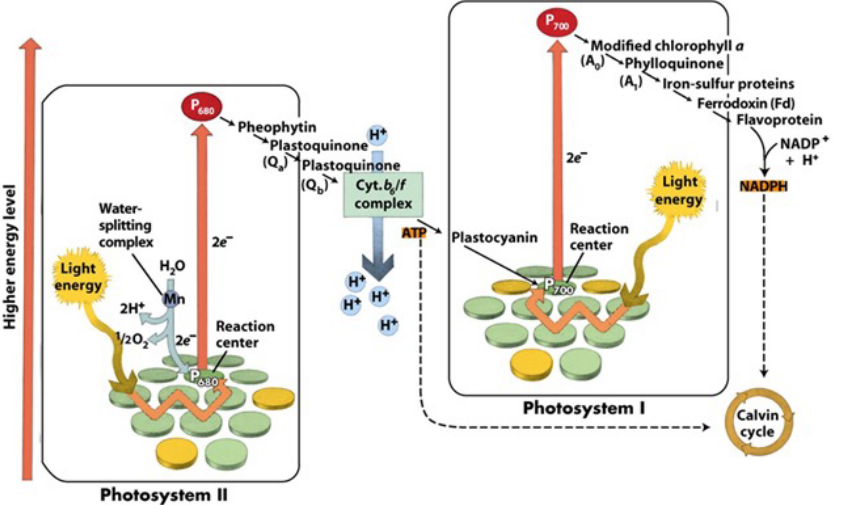
Figure 12.2: Phototrophy
In anoxygenic photosynthesis, neither photosystem I or II is able to oxidize H2O to O2.
In oxygenic photosynthesis, the following reaction is possible:
\[\begin{equation} CO_2 + H_2O \rightarrow C_6H_{12}O_6 + O_2 \end{equation}\]
12.2.1 Photosynthesis
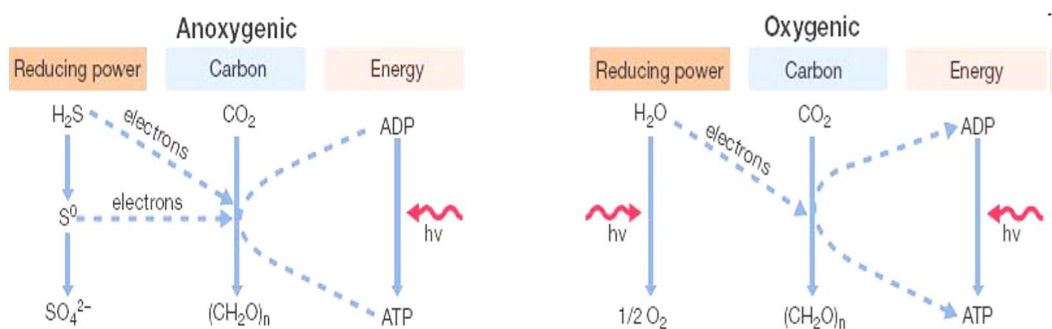
Figure 12.3: Two Types of Phototrophy
Oxygenic photosynthesis not only uses PSI and PSII, but also generates considerably more energy (36 molecules of ATP) compared to anoxygenic photosynthesis and generates O2 and electrons from water. Oxygenic phototrophs are cyanobacteria.
Anoxygenic photosynthesis, on the contrary, predates oxygenic photosynthesis; anoxygenic phototrophs include green sulfur, green non-sulfur, purple sulfur, and non-purple sulfur bacteria. However, the electrons that are present in the electron transport chain (in anoxygenic photosynthesis) do not come from water.
The machinery for anoxygenic photosynthesis is also the building block for oxygenic photosynthesis.
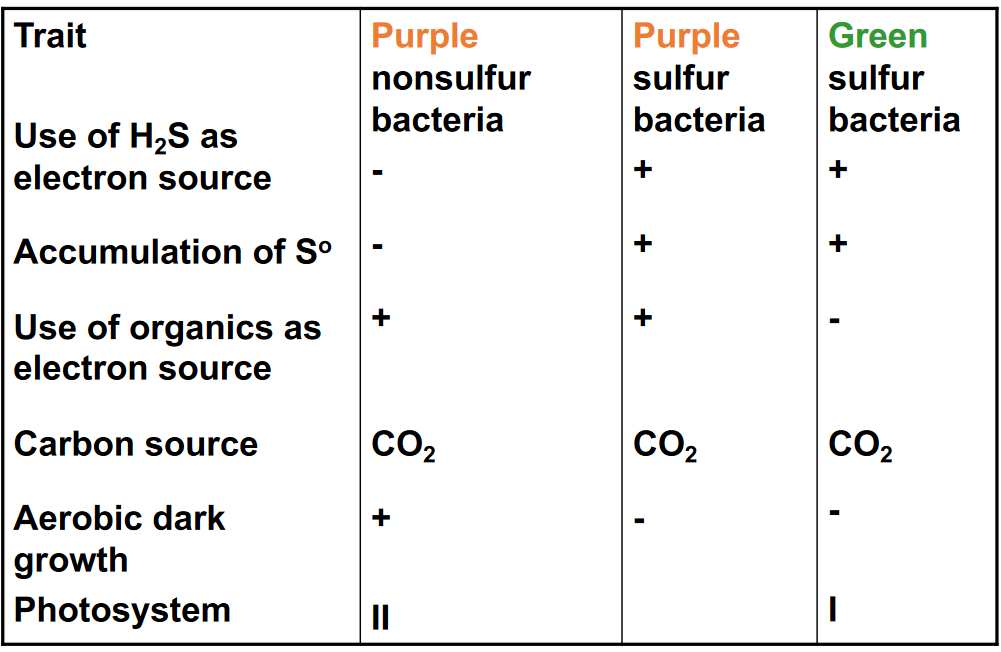
Figure 12.4: Anoxygenic Bacterial Photosynthesis
12.3 Chemotrophy
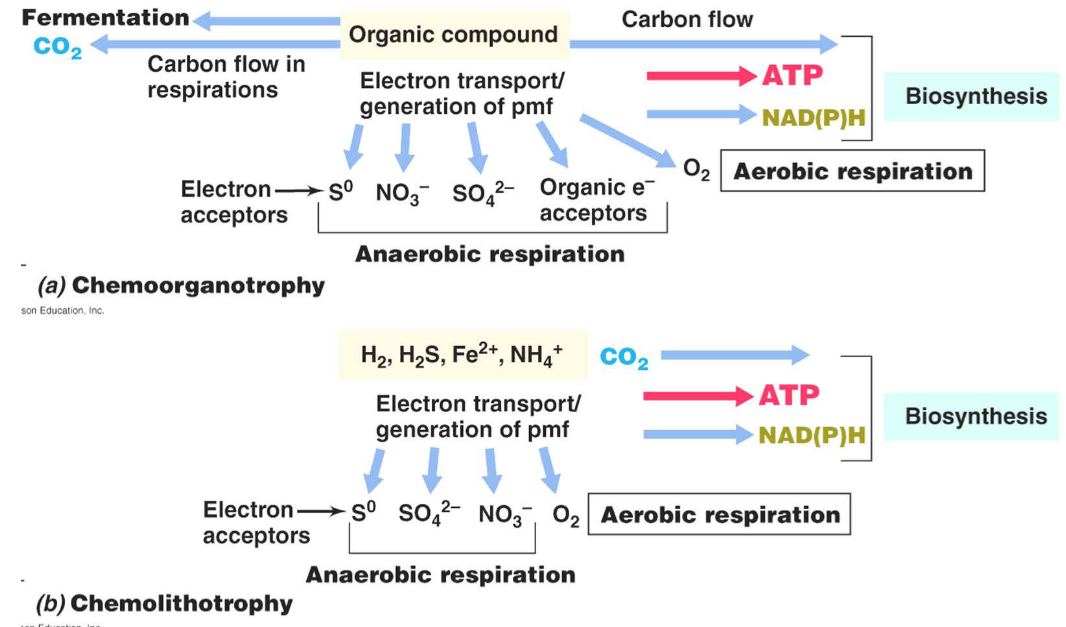
Figure 12.5: Types of Chemotrophs
A chemoheterotroph is an organism that uses energy and carbon from organic substrates.
Chemolithotrophs obtain their energy from oxidizing inorganic compounds (e.g., NH3, Fe2+, H2S, etc).
Photoheterotrophs obtain their energy from light and their carbon from organic sources.
Photoautotrophs also obtain their energy from light, but their carbon from CO2.
12.3.1 Anaerobic respiration
Such respiration uses inorganic electron acceptors (other than O2) in oxidative phosphorylation.
Consequently, there is also a truncated respiratory electron transport system and a lesser amount of energy generation than aerobic respiration.
Anaerobic respiration is also only carried out by prokaryotes:
- Obligate anaerobes are organisms that cannot respire in the presence of O2.
- Facultative anaerobes are organisms that can respire in the presence or in the absence of O2,
12.3.2 Sulfates
12.3.2.1 Sulfate reduction
\[\begin{align} SO_4^{2-} + 2H^+ + 2e^- &\rightarrow SO_3^{2-} + H_2O &\text{ (1 ATP)} \\ SO_3^{2-} + 6H^+ + 6e^- &\rightarrow S^{2-} + 3H_2O &\text{ (3 ATP)} \end{align}\]
Sulfate-reducing bacteria have short electron transport chains - this is because very little energy is available (relative to O2).
Sulfidogens are microbes that produce H2S. Obligate anaerobes are common in soil and marine environments. These microbes are also very diverse (e.g., Desulfovibrio).
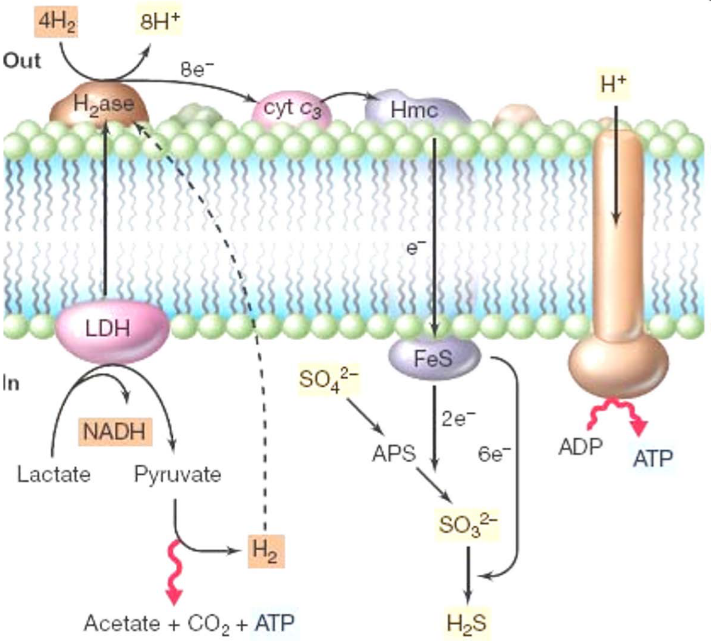
Figure 12.6: Sulfate Reductions
Sulfate reduction can look something like the above graphic.
12.3.2.2 Sulfate oxidation
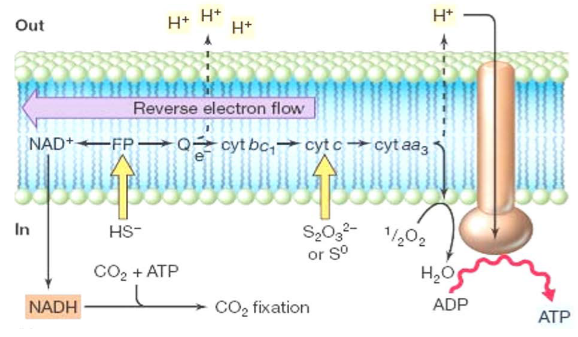
Figure 12.7: Sulfate Oxidation in Bacteria
In sulfate oxidizers, the electron transport chain can be reversed. This is because HS-, S2O32-, and S0 are weaker electron donors than NAD / NADH.
A proton motive force is also formed at the quinone (Q) and the terminal oxidase (cytochrome - cyt).
12.3.3 Nitrogen
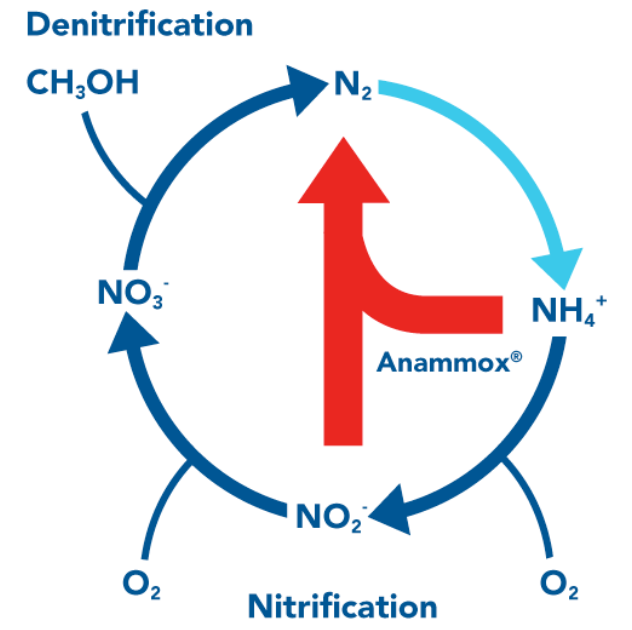
Figure 12.8: Nitrogen Cycle
There are many steps to the nitrogen cycle (shown above)
12.3.4 Denitrification
There are four main reactions to be wary of:
\[\begin{align} NO_3^- + 2H^+ + 2e^- &\rightarrow NO_2^- + H_2O &\text{ (2 ATP)} \\ 2NO_2^- + 2H^+ + 2e^- &\rightarrow 2NO + 2OH^- &\text{ (2 ATP)} \\ 2NO + 2H^+ + 2e^- &\rightarrow N_2O + H_2O &\text{ (2 ATP)} \\ N_2O + 2H^+ + 2e^- &\rightarrow N_2 + H_2O &\text{ (2 ATP)} \end{align}\]
Nitrate respiration is carried out by E. coli and many other bacteria. Denitrification is the sequential reduction of NO2 to nitrogen gases (e.g., N2 and N2O). Nitroux oxide (i.e., N2O) is a potent greenhouse gas made from denitrification in agricultural soil (it is also the main constituent in laughing gas).
12.3.4.1 Nitrogen fixation
Life depends on the quick cycling of nitrogen into biologically available forms.
Nitrogen fixation reduces dinitrogen gas (i.e., N2) to ammonia (i.e., NH3) using the enzyme nitrogenase (which is very sensitive to O2).
Nitrogen fixation is also a two-step process - it requires dinitrogenase and dinitrogenase reductase. Only prokaryotes are able to carry out nitrogen fixation: symbiotic bacteria (i.e., Rhizobia), free-living anaerobes (Clostridium), free-living aerobes (i.e., Azotobacter), and those that are able to perform cell differentiation (i.e., Cyanobacteria).
12.3.4.1.1 Nitrogen fixing bacteria
knitr::include_graphics("_book/_main_files/figure-html/rhizobia.png")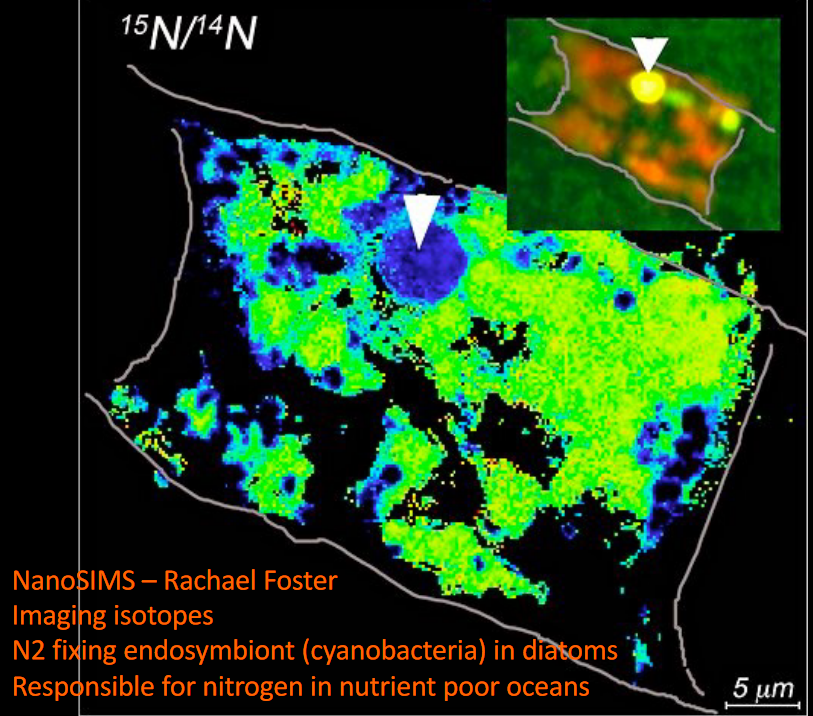
Figure 12.9: Rhizobia in Eukaryotes
Rhizobia are symbiotes with terrestrial plants - eukaryotes are unable to metabolize atmospheric nitrogen, so Rhizobia are essential to fix atmospheric nitrogen.
Rhizobia also induces cell differentiation in the host plant to create a niche. Root nodules have also reduced oxygen as nitrogen fixation is an anaerobic process.
12.3.4.1.2 Aerobic N2 fixation
Azotobacter vinelandii is able to perform aerobic N2 fixation.
Under hypoxic conditions, the bacterium does not produce a slime layer. Otherwise, there is a large slime layer under hyperoxic conditions.
Slime retards the diffusion of O2 into the cell, hence also implying that nitrogenase will not be inactivated.
12.4 Metabolic Interactions
Microbes have a much more diverse metabolism than eukaryotes as they can use light and inorganic chemicals as energy sources. Eukaryotes are driven by mitochondria (an alpha proteobacteria) - an organelle that is only able to use organic carbon for energy.
However, bacteria and archaea often have their own source of electron acceptors and donors from other microbes, hence allowing them to live symbiosis with other microbes in a condition called metabolic consortia.
For instance, one microbe may take the waste products of another and use that as a source of electrons, hence also degrading the electron source (i.e., reducing agent) until it becomes a better electron dump (i.e., oxidizing agent).
12.4.1 Heterocysts
Cyanobacteria are able to perform nitrogen fixation in aquatic systems - this fixation is also performed in specialized cells called heterocysts.
Heterocysts have a thickened cell wall and have no chlorophyll (and hence unable to conduct photosynthesis). There is also a reduced amount of nitrogen that is transported from the heterocyst to vegetative cells and sugar transported from vegetative cells to heterocysts.
Sugars are also supplied in excess so that they leak out of the cell, hence attracting heterotrophic bacteria that consume O2 gas and reduces oxygen.
12.4.2 Metabolic consortia

Figure 12.10: Metabolic Consortium
Some metabolisms are performed by a consortium of bacteria; however, some steps may need to be physically separated from other steps when possible. There is also an adequate amount of energy from a single step in the metabolism.
When metabolites are small and soluble, they diffuse. Colonies that are barely visible and can be re-grown, but this is difficult and and contributes to the low number of microbial species in culture.
Substances that don’t diffuse are usually:
- Large (e.g., polymers)
- Insoluble (e.g., hydrophobic)