Chapter 11 Renal Physiology I
The main homeostatic factors for generating urine are:
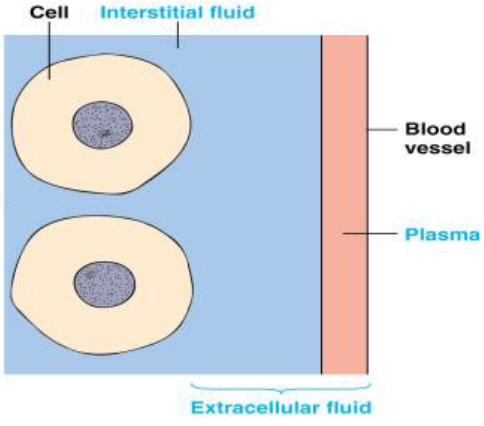
Figure 11.1: Extracellular Fluid (i.e., ECF)
- Nutrient concentrations
- Waste product concentrations
- O2 and CO2 concentrations
- pH
- Ion concentrations
- Temperature
- Volume and pressure
11.1 Overview of the Renal System
The renal system regulates homeostasis. More specifically though, it regulates the volume of the ECF and also the quantity of most ECF ions: including Na+, K+, Ca2+, PO4-, and HCO3-.
Furthermore, the renal system also excretes waste products (e.g., urea, uric acid, creatinine, etc.) and foreign products (e.g., drugs and pesticides).
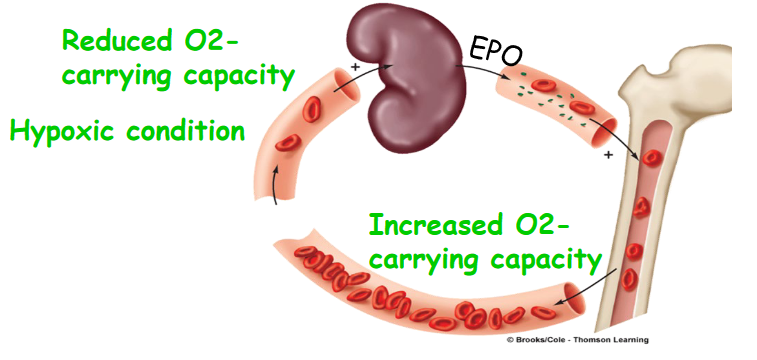
Figure 11.2: EPO
The renal system also regulates the pH of the blood by adjusting urinary acid excretion and produces (e.g., renin and erythroprotein [EPO]) and activates hormones (e.g., T3 and T4).
Also, study the following graphic:
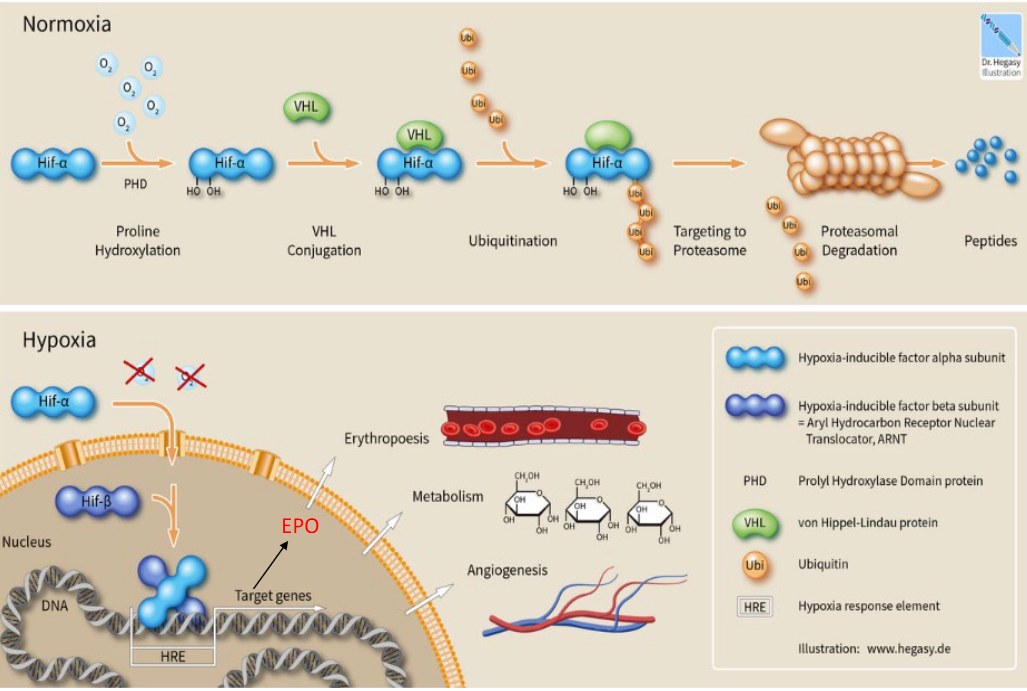
Figure 11.3: Cellular Response to Normoxic and Anoxic Conditions
Erythroprotein is a cytokine produced by the kidneys that enhance the production of red blood cells. This is especially crucial during a person’s adaptation to higher altitudes (especially since there is a lack of O2 to go around).
11.2 The Kidneys
Observe the structure of the kidneys:
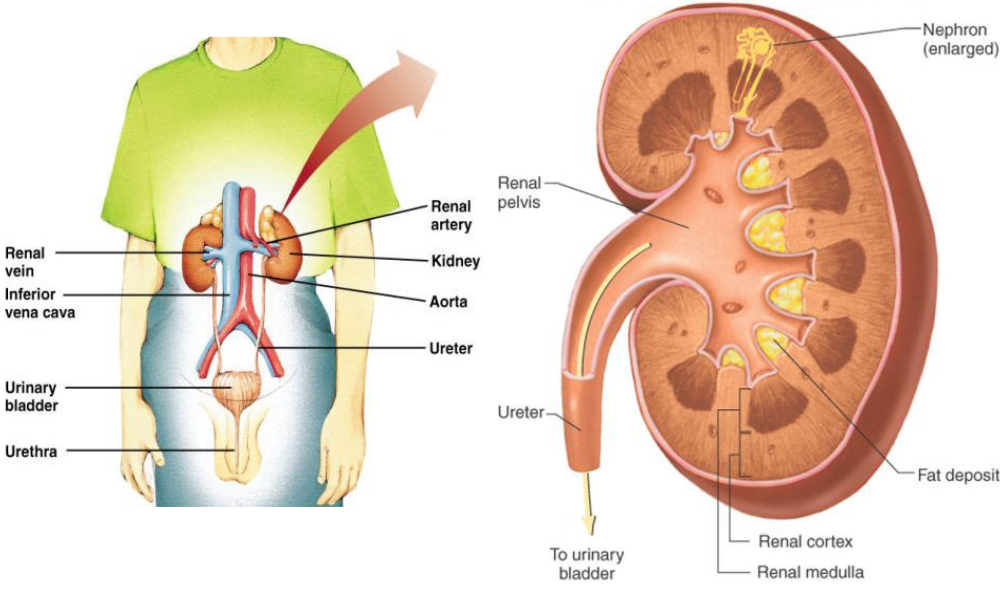
Figure 11.4: Structure of the Kidneys
11.2.1 Nephrons
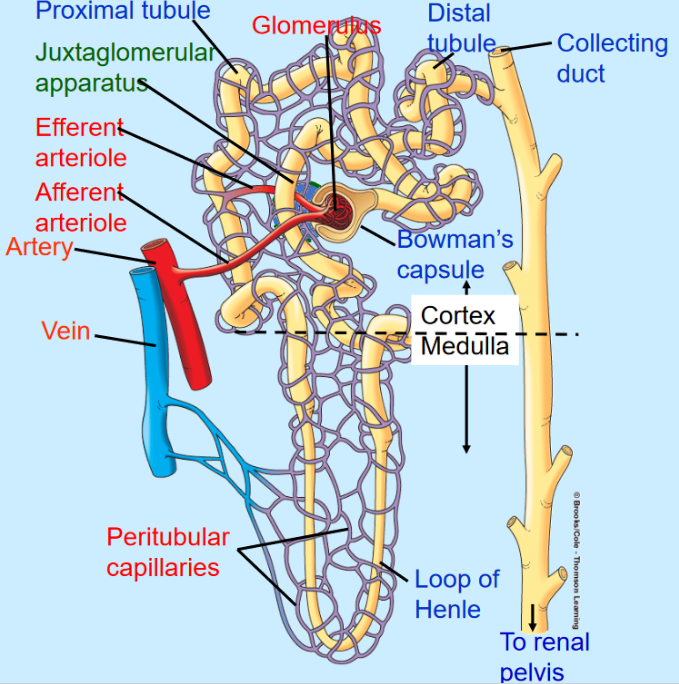
Figure 11.5: Structure of a Nephron
Nephrons are the basic, functional unit of the kidneys. A nephron has two components:
The tubular component: this includes the Bowman’s capsule, the proximal tubule, the loop of Henle, and the distal and proximal collecting tubes.
There is also the vascular component. The afferent arteriole in this component delivers blood into the glomerulus: a capillary network that produces filtrate that enters the urinary tubules. The efferent arteriole delivers blood from the glomeruli to peritubular capillaries: vessels that deliver blood to renal tissue and are important in the exchange of waste products between tubular system and blood in the production of urine.
11.3 Basic Renal Processes
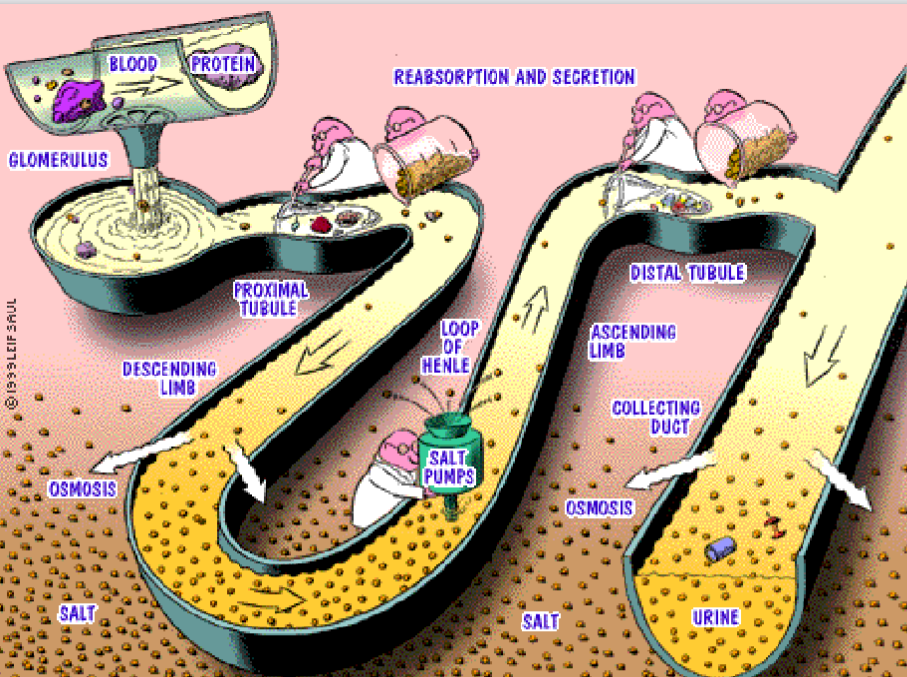
Figure 11.7: Processes in the Nephrons
The three basic renal processes are:
Glomerular filtration
This is the filtration of protein-free plasma through the glomerular capillaries into the Bowman’s capsule.
Tubular reabsorption
This is the return of most solutes and H2O from the urine filtrate into the peritubular capillaries.
Tubular secretion
This is the selective transfer of substances from the peritubular capillary blood into the tubular lumen.
11.3.1 Glomerular filtration
Filtration by the glomerulus includes the following three components:
- Glomerular filtration membrane
- Filtration pressure
- Glomerular filtration rates and regulation
11.3.1.1 Glomerular filtration membrane
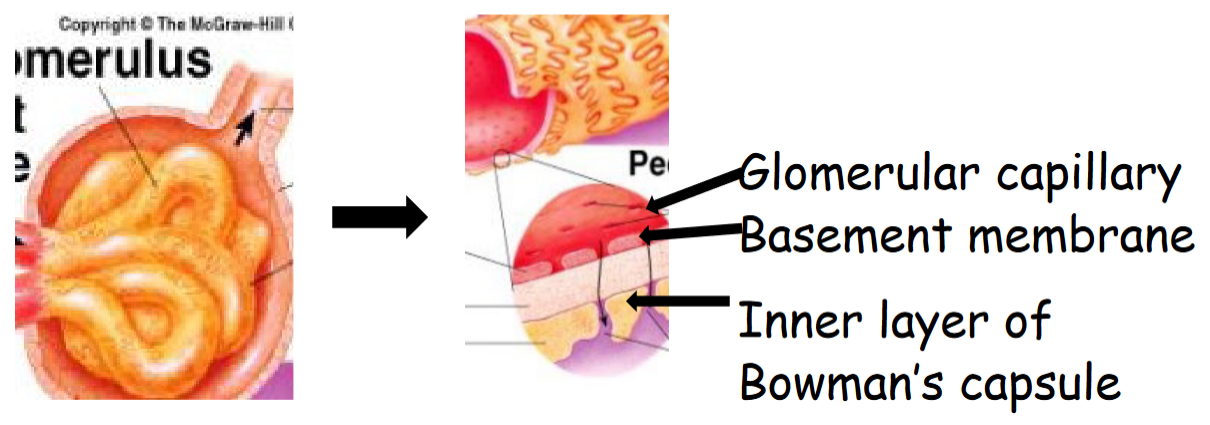
Figure 11.8: Glomerular filtration Membrane Structure
The glomerular capillary have pores that are 100 times more permeable than pores elsewhere.
The basement membrane is made up of negatively-charged collagen and glycoproteins - this prevents the leakage of negatively-charged plasma proteins. Post-streptococcal Glomerulonephritis is caused by the bacterium depositing proteins on the glomerulus’ membrane.
The inner layer of the Bowman’s capsule contains octopus-like cells with podocytes with intercalcating foot processes. Narrow slits between the foot processes are used for filtration.
11.3.1.2 Filtration pressures
Pressure varies depending on the tubular component being examined; the:
- Glomerular capillary pressure is around 55 mm Hg. It is higher than the capillary pressure elsewhere - this because the diameter of the afferent arteriole is wider than the efferent arteriole.
- Plasma-colloid osmotic pressure is around 30 mm Hg. This pressure occurs between the Bowman’s capsule and the capillaries. The pressure opposes filtration.
- Bowman’s capsule hydrostatic pressure is around 15 mm Hg, This pressure also opposes filtration.
Hence, the net filtration is (50 - 15 - 30) mm Hg = 10 mm Hg. The filtration pressure is influenced by glomerular capillary pressure - this is determined via the diameter of the afferent and efferent arterioles!
11.3.1.3 Glomerular filtrate rate (GFR)
The GFR is denoted by:
GFR=Kf×(net filtration pressure)
Note that Kf is determined by the surface area of the glomerulus and glomerulus’ membrane permeability.
20% of the plasma entering the glomeruli is filtered; about 180 ml of blood is filtered per day and about 125 ml per minute.
11.3.1.4 Regulation of filtration pressure
Autoregulation prevents a change in the filtration pressure. This mechanism maintains the GFR between (80 and 180 mm Hg).
Sympathetic regulation changes the filtration pressure in response to a decrease in blood pressure via the baroreceptor reflex.
Epinephrine and angiotensin II from the adrenal glands also help to regulate the filtration pressure.
11.3.1.4.1 Autoregulation
There are two mechanisms for this:
The myogenic mechanism occurs when the arterioles constrict when stretched because of increased pressure. Consequently, when the blood pressure increases, the smooth muscle cells of the wall are stretched and respond by being contracted (hence resulting in little change in flow). When blood pressure falls, the smooth muscle cells of the wall relax and allow for the continued flow of blood.
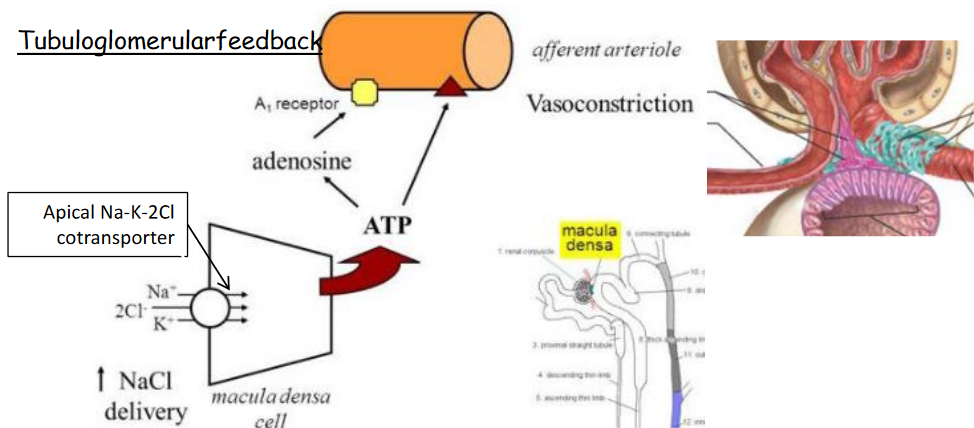
Figure 11.9: Tubuloglomerular feedback
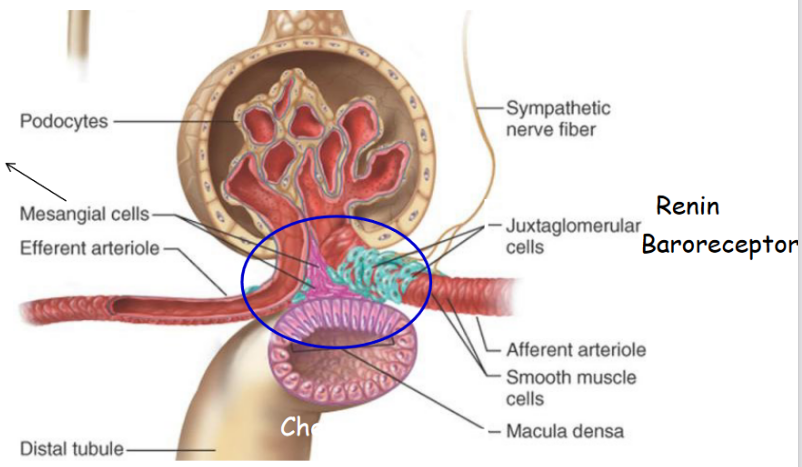
In tubuloglomerular feedback, a rise in NaCl concentration in the fluid of the distal tubule causes the Macula densa cells to produce adenosine - this causes afferent arterioles to vasoconstrict. Adenosine also inhibits the release of renin.
Similarly, a decrease in the concentration of NaCl also causes Macula densa cells to release PGE2, hence causing vasodilation and encouraging renin release.
11.3.1.4.2 Sympathetic regulations
Baroreceptors are able to detect a significant fall in blood pressure - when it does, this leads to the activation of the sympathetic nervous system and hence, the vasoconstriction of afferent arterioles.
Sympathetic regulations are able to override autoregulation; there is also no evidence for parasympathetic innervation in the kidney.
Such a regulation also decreases the flow of blood through the kidneys, hence also decreasing the GFR. When blood pressure falls too much, the sympathetic nerves may also stimulate the release of renin (which - through a protein-cleaving reaction - generates angiotensin II). Angiotensin II, in turn, also stimulates aldosterone production to augment blood volume through the retention of Na+ and water.
11.3.2 Tubular reabsorption
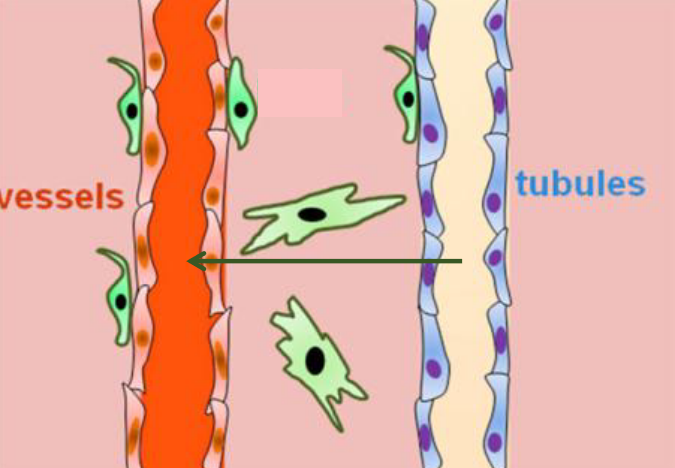
Figure 11.10: Tubular Reabsorption
Like mentioned previously, most solutes and H2O from the urine filtrate is returned into the bloodstream.
Funnily enough, of the 180 L of blood that is filtered each day, only about 1 to 2 liters of urine is excreted by the body each day. A small reduction in the reabsorption can lead to a large increase of urine volume.
That said, there are also barriers for tubular reabsorption:
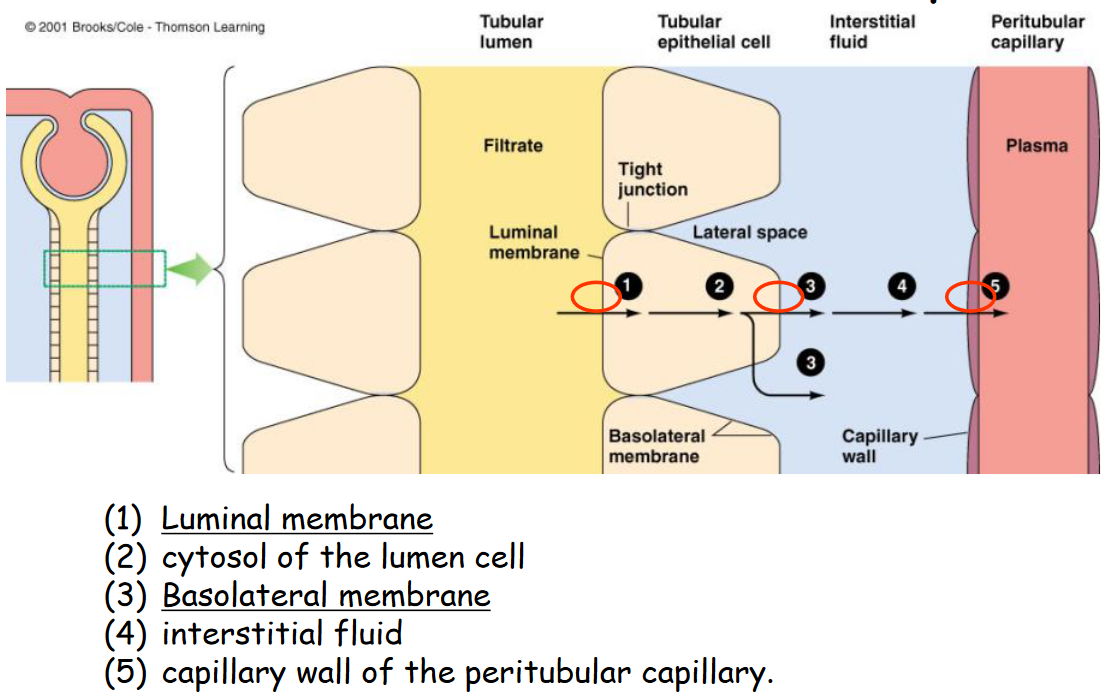
Figure 11.11: Barriers to Tubular Reabsorption
And of the minerals that are reabsorbed, the composition of the reabsorbed minerals are:
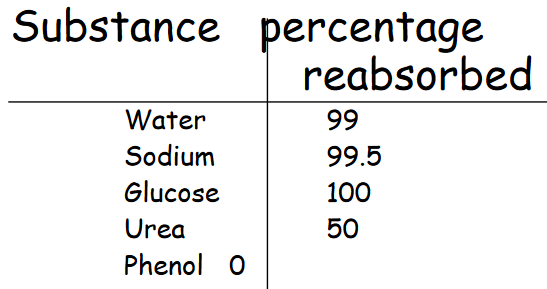
Figure 11.12: Reabsorbed Substances
11.3.2.1 Mechanism of sodium reabsorption
Observe the following graphic:
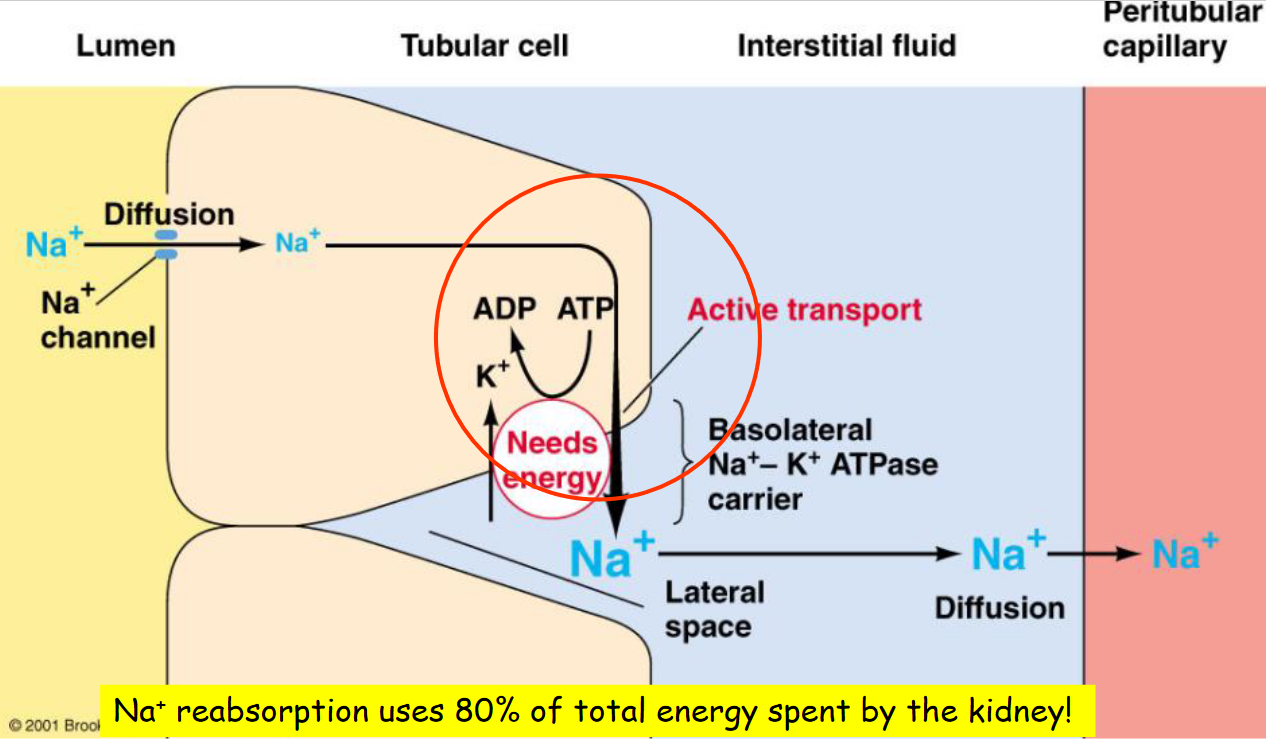
Figure 11.13: Sodium Reabsorption
Na+ reabsorption in the proximal tubule (about 67%) is crucial for the re-absorption of glucose, amino acid, urea, H2O, and Cl- ions. Note that the descending loop of Henle is not permeable to Na+.
Only the ascending loop of Henle is permeable to Na+ - critical for water elimination or conservation.
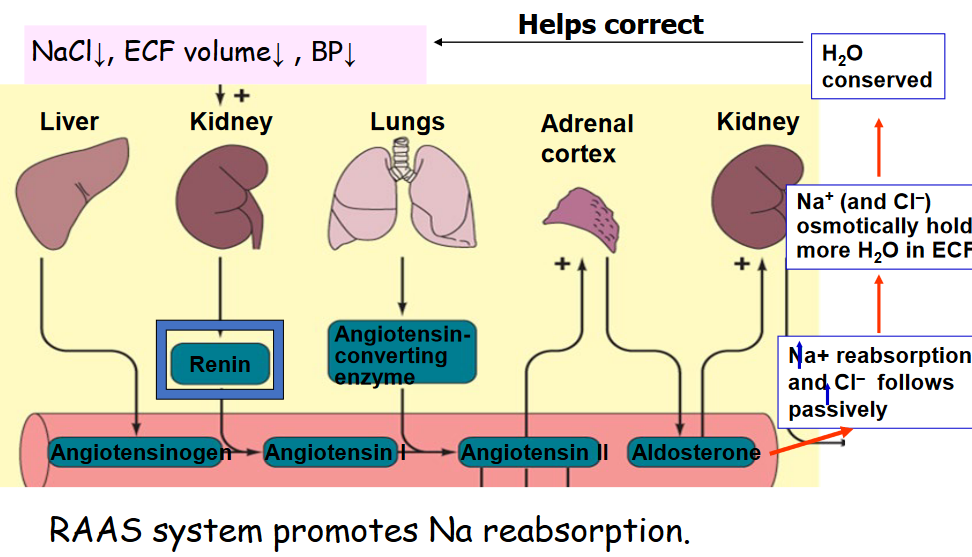
Figure 11.14: RAAS Regulation of Sodium Reabsorption
Na+ reabsorption in the distal and collecting tubules are also subject to hormonal control. About 8% of all filtered sodium depends on aldosterone for re-absorption, and in the presence of maximal aldosterone secretion, all filtered Na+ is re-absorbed through Na channels and Na+ / K+ ATPase.
11.3.2.1.1 Neural regulation of sodium reabsorption
A fall in blood pressure triggers an increase in the activity of the sympathetic nerves that innervate the granular cells.
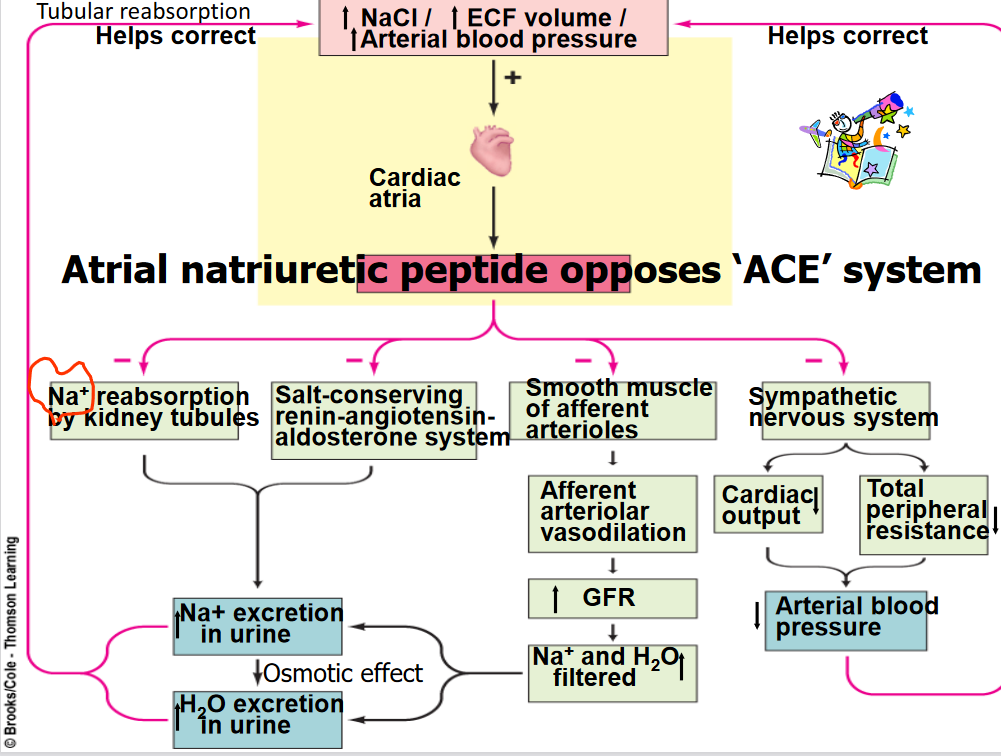
Figure 11.15: RAAS Activation of Sodium Reabsorption
Both the systemic and local input will lead to the release of renin, hence activating the Renin-angiotensin-aldosterone system.
11.3.3 Reabsorbing H2O (tubular reabsorption)
H2O is reabsorbed by osmosis. 80% of H2O is reabsorbed in the proximal tubules and the descending limb in the loop of Henle. Water channels in these tubular components are always open.
The remaining 20% H2O is absorbed by the distal tubules and collecting ducts. However, this depends on the insertion of aquaporin 2 under the regulation of ADH (i.e., vasopressin).
11.3.3.1 Aquaporin 1
There are six trans-membrane domains:
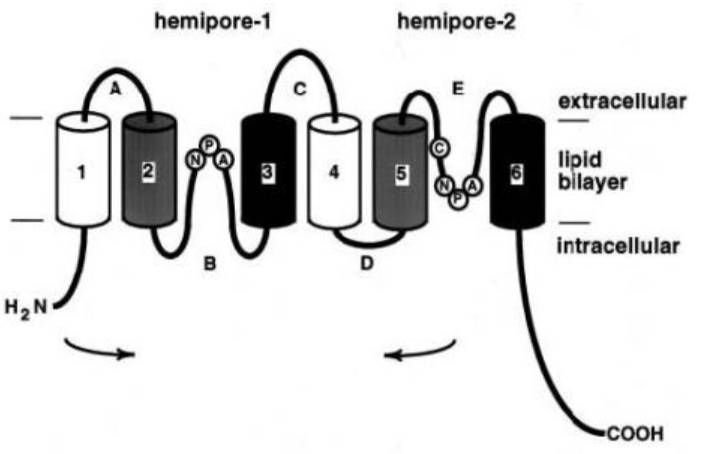
Figure 11.16: Transmembrane Domains of Aquaporin 1
These trans-membrane domains exist as a tetramer where each subunit has its own water pore. While loops B and E are juxtaposed to form a water channel, HgCl binds to a cysteine residue in the E loop to close the channel.
Shown below is the structure of aquaporin 1:
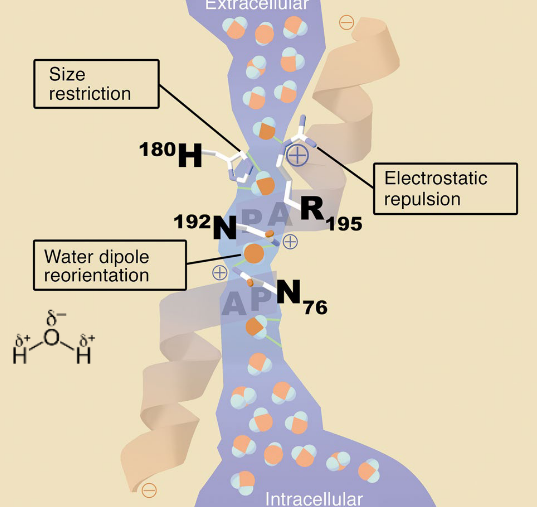
Figure 11.17: Aquaporin 1 Subunit
Also, here is the mechanism by which vasopressin encourages water retention:
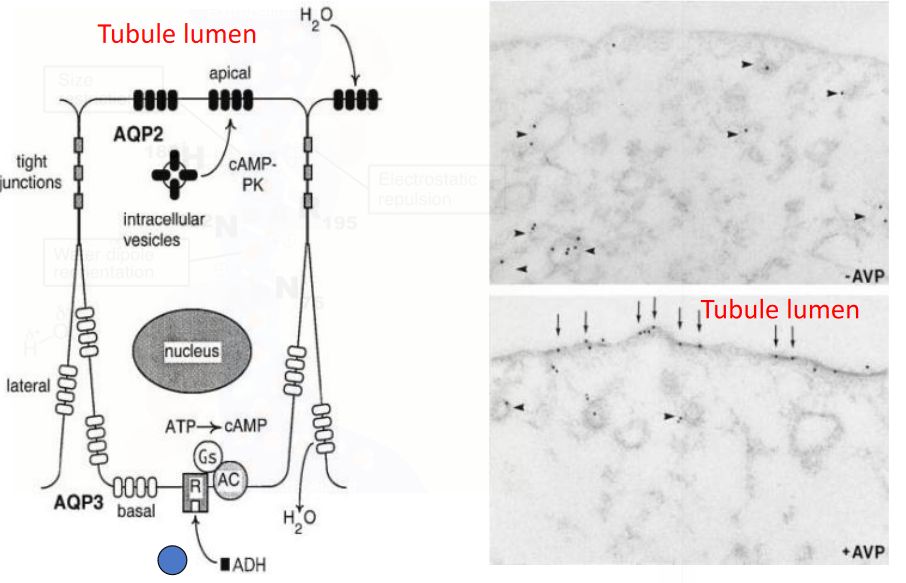
Figure 11.18: Vasopressin Stimulates Water Retention
11.3.4 Glucose Reabsorption
Glucose is reabsorbed by Na+-dependent secondary active transport using the Na+-glucose co-transporter in the proximal tubule.
Note that the absorption of glucose has a maximum rate Tm (in minutes) due to the limited number of carrier proteins. The Tm for glucose is 375 mg / minute.
Hence, glucose is excreted when the filtered load is greater than the Tm (i.e., the plasma concentration of the substance × GFR)
The renal threshold is the plasma concentration at which the Tm of a substrate is reached - It is greater than or equal to 300 mg / 100 ml of blood for glucose.
In theory, diabetic patients have glucose concentrations that are three times that of a normal patient’s. Because of this, glucose in the bloodstream may spill over into the urine at 180 mg / 100 ml.
11.3.5 Reabsorbing Urea
Urea reabsorption does not exhibit the tubular maximum - nearly half of the filtered urea is reabsorbed in the proximal tubule.
Medullary collecting ducts are also highly permeable to urea as urea is the most concentrated product in this segment.
Note that reabsorbing urea creates a **hyperosmotic medullary interstitium* - this is a crucial step in making concentrated urine. ADH increases the permeability of the medullary collecting ducts via the urea transporters.