Chapter 12 Nucleotide and Nucleic Acid Metabolism
Nucleic acids are the chemical carriers of an organism’s genetic information. There are two classes of nucleic acids: DNA (stores and transfers genetic information) and RNA (has a broader range of functions - some classes of RNA are found in cells)
There are three kinds of RNA examined in BS2003:
- Ribosomal RNAs (i.e., rRNAs) - these are components of ribosomes: complexes that carry out protein synthesis.
- Messenger RNAs (i.e., mRNAs) - these carry genetic information from at least one gene to a ribosome.
- Transfer RNAs (i.e., tRNAs) - these are adapter molecules that translate the information in mRNA into a specific sequence of amino acids.
12.1 What are Nucleic Acids?
Nucleic acids are polynucleotides: each nucleotide three parts:
- A five-membered cyclic monosaccharide
- A phosphate group
- A nitogen-containing cyclic compound
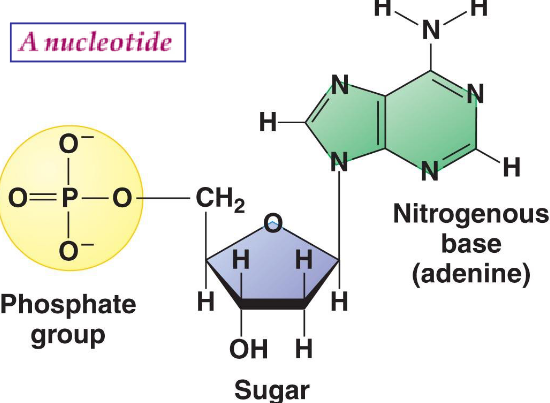
Figure 12.1: Parts of a Nucleotide
Note that successive nucleotides of RNA and DNA are bound via phosphodiester linkages. The 5-phosphate group of one nucleotide unit is joined to the next 3-hydroxyl group of the next nucleotide. All phosphodiester linkages have the same orientation, hence giving each linear nucleic acid strand a specific polarity:
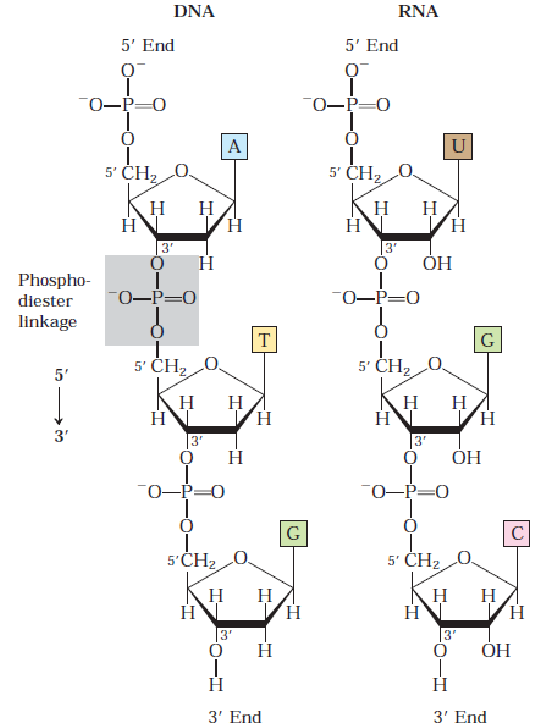
Figure 12.2: Nucleotide Linkages in DNA
12.1.1 Nucleosides
Note that all nucleosides are named by adding the suffix -idine to a pyrimidine (i.e., a monocyclic structure) and the suffix -osine to a purine (i.e., a polycyclic structure).
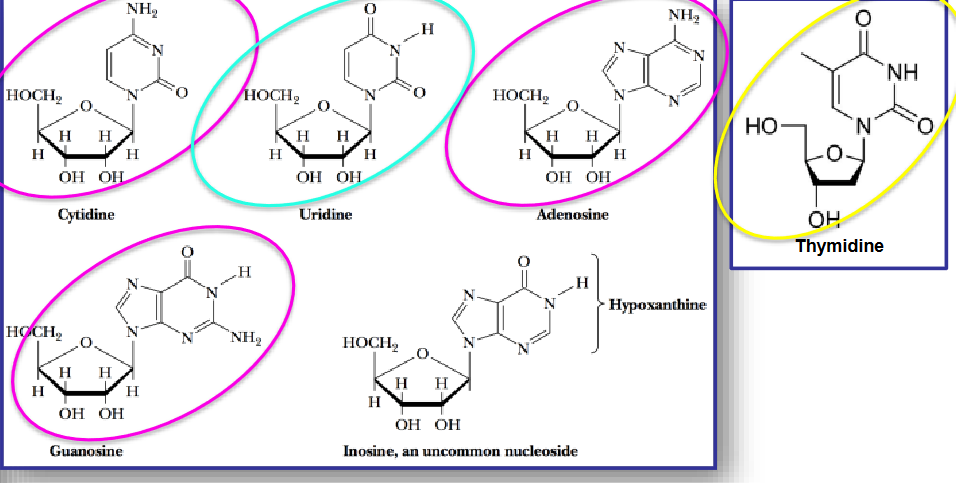
Figure 12.3: Nucleoside Structures
The most common nucleosides are adenine, thymidine, cytodine, guanidine, and uridine. The pyrimidines are cytodine thymidine, and uridine - the purines are adenosine and guanine.
In mammals, adenosine functions as an autacoid or a “local hormone.” Caffeine is an alkaloid:
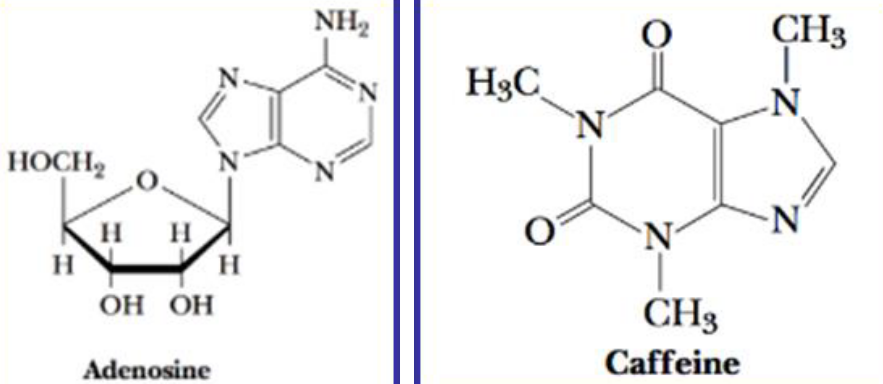
Figure 12.4: Structure of Caffeine and Adenosine
Caffeine promotes wakefulness by blocking the interaction of extracellular adenosine with its neuronal receptors.
Autacoids also perform a wide variety of functions:
knitr::include_graphics("_book/_main_files/figure-html/autocoidfunction.png")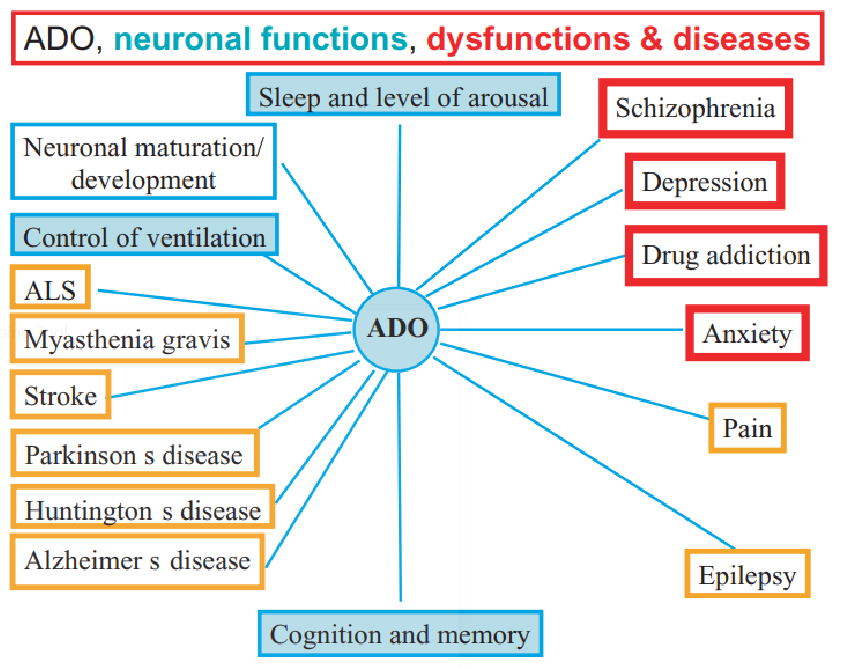
Figure 12.5: Autocoid Functions
12.2 Nucleotide Roles in Metabolism
Nucleotides are…
- Precursors of DNA and RNA synthesis
- Carriers of chemical energy (e.g., ATP)
- Cofactors (e.g., NAD, FAD, etc.)
- Activated intermediates (e.g., UDP-glucose, etc.)
- Second messengers (e.g., cAMP, cGMP, etc.)
12.2.1 Chemical energy from nucleotides
ATP, AMP, and ADP are needed for energy metabolism, pumping material across the cell membrane, and muscle contraction.
UTP is needed for sugar interconversion and oligosaccharide synthesis.
GTP is needed to form GDP-activated sugars of complex carbohydrates. GTP also interacts with G proteins and forms a peptide chain.
Cyclic AMP is needed for metabolic control and modifies enzyme properties. Cyclic GMP is needed for vision and metabolic control. (p)ppGpp is an alarmone produced by bacteria in response to a reduction in protein synthesis during amino acid starvation - rRNA and tRNA synthesis is also inhibited here.
Adenine nucleotides are components of many enzyme cofactors. While adenosine does not actively participate in any enzymatic reactions, removing the adenine portion of the cofactor drastically reduces the cofactor’s activities.
12.3 Biosynthesizing Purines and Pyrimidines
There are two kinds of pathways:
- De novo pathways use amino acids, ribose-5-phosphate, CO2, and NH3 to make purines and pyrimidines.
- Salvage pathways recycle free bases and nucleotides released from nucleic acid decomposition.
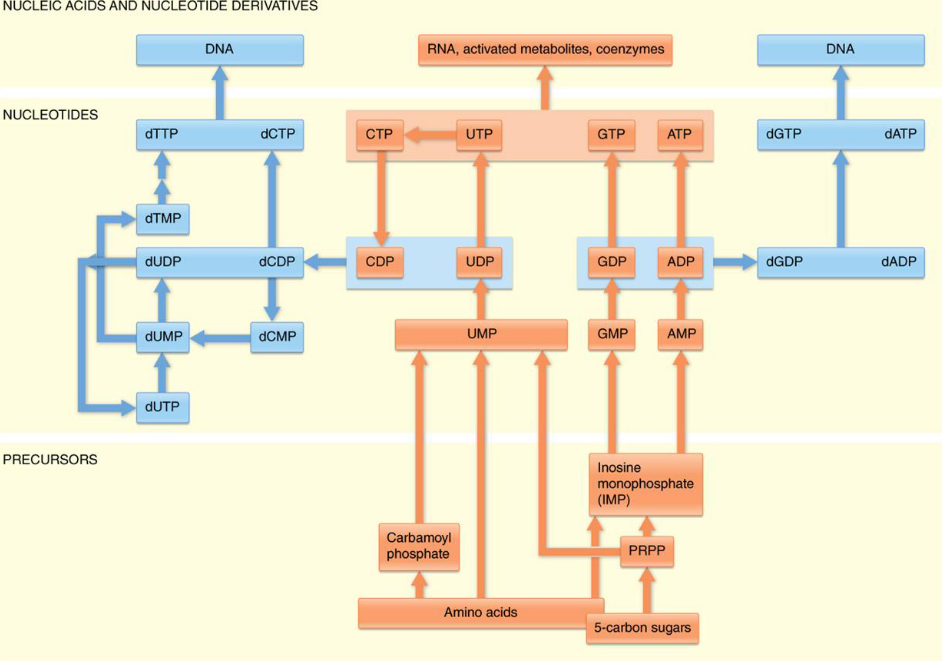
Figure 12.6: De novo Pathways
The three most important precursors in the above biosynthesis reactions are:
- PRPP
- Carbamoyl phosphate
- Glycine, aspartate, and glutamine
Note that purine rings are built atom-by-atom on the ribose base. The ribose base is attached after the pyrimidine ring is formed.
12.3.1 Making purines
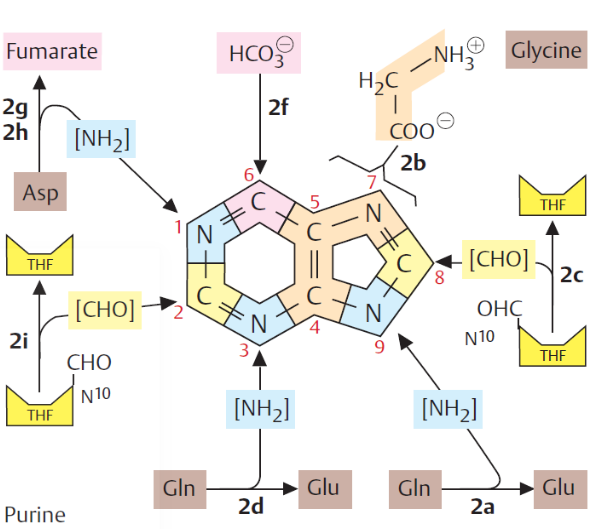
Figure 12.7: Atom Sources of a Purine Ring
In 1949, it was discovered that birds excrete excess nitrogen in the form of uric acid. From this, it was also discovered that the nine atoms of a purine ring system are contributed by different molecules.
Nevertheless, most de novo synthesis occur in the liver; the nitrogenous bases and nucleosides are then transported to other tissues via the erythrocytes (i.e., the red blood cells).
Purines are made as ribonucleotides - the initial purine that is synthesized is inosine monophosphate (i.e., IMP). AMP and GMP are derived from IMP in two-step reaction pathways.
12.3.1.1 PRPP
Figure 12.8: PRPP
5-Phospho-\(\alpha\)-D-ribosyl 1-pyrophosphate (better known as PRPP) is a key intermediate in the de novo synthesis of purine and pyrimidine nucleotides.
12.3.1.2 Synthesizing IMP
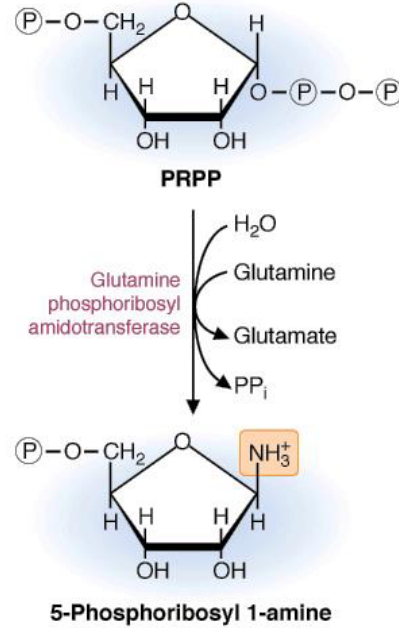
Figure 12.9: IMP Synthesis
There are several steps in synthesizing IMP:
The first step of purine biosynthesis is the committed step of the pathway. The purine base is built on top of this moiety; however, because 5-Phosphoribosyl is highly unstable, the availability of PRPP is a major determinant of the rate of the above reaction.
In the second step of purine biosynthesis, there is an addition of three carbons from glycine:
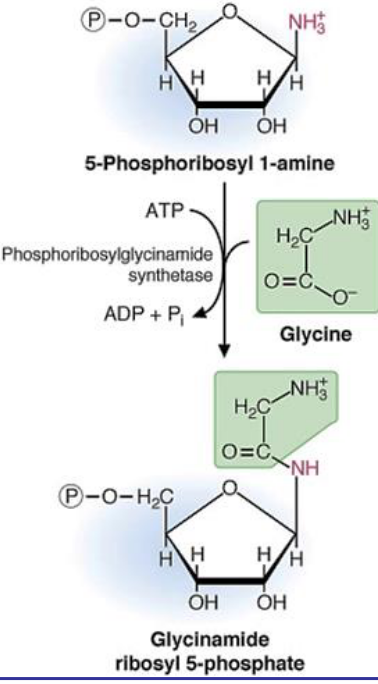
Figure 12.10: Second Step of IMP Biosynthesis
Note that the ATP above is needed for the condensation of the glycine carboxylic acid group with the 1’ amino group of phosphoriboyl 1-amine.
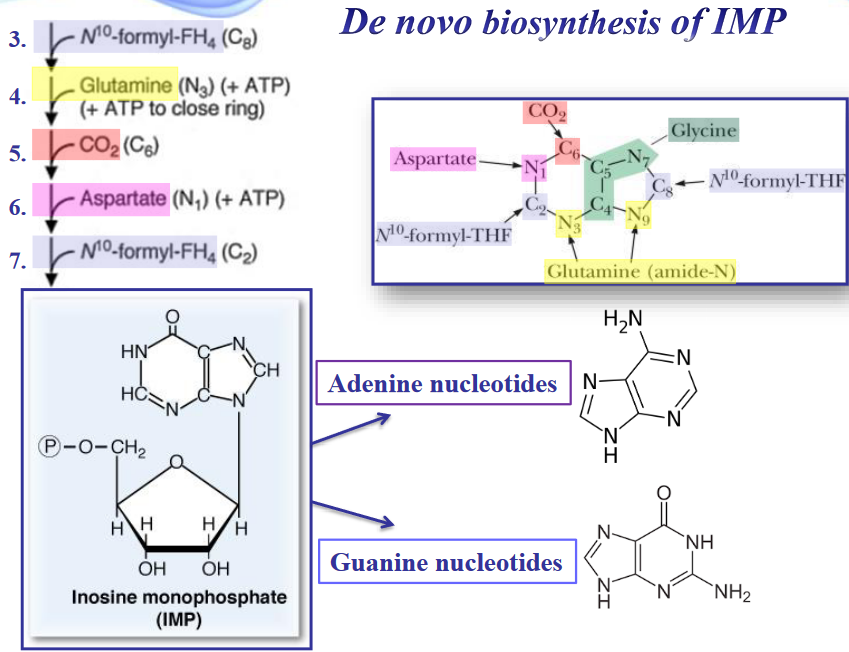
Figure 12.11: Sources of Carbons in Purine Synthesis
12.3.2 ATP-dependent kinases and nucleoside (di / tri)-phosphates
The molecules AMP and GMP are converted by successive phosphorylation into their metabolically prominent forms: ATP and GTP.
The first two phosphorylations are carried out by adenylate kinase and guanylate kinase:
\[\begin{align} \text{Adeylate Kinase: } AMP + ATP &\rightarrow 2 ADP \\ \text{Guanylate Kinase: } GMP + ATP &\rightarrow GDP + ADP \end{align}\]
The above kinases also act on deoxynucleotide monophosphates to yield dADP or dGDP. Note that oxidative phosphorylation is usually responsible for the conversion of ADP into ATP.
ATP can also participate in the following reaction - one that is catalyzed by nucleoside diphosphate kinase:
\[\begin{equation} GDP + ADP \rightarrow GTP + ADP \end{equation}\]
12.4 Salvage Pathways
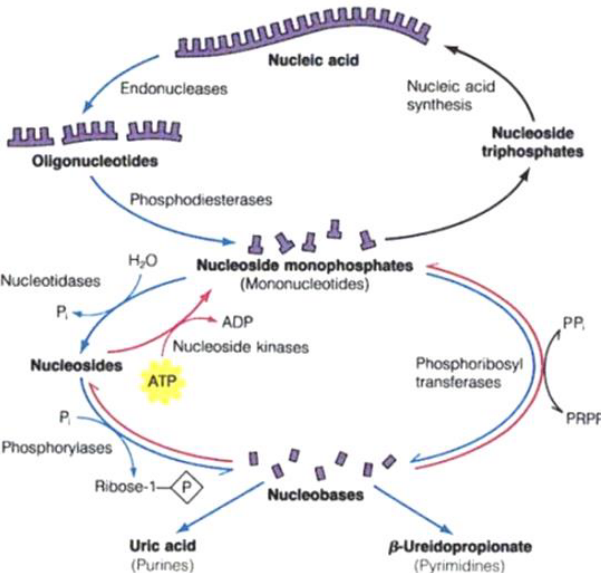
Figure 12.12: Salvage Pathways
Catabolic reactions in the above graphic are shown in blue; nucleotide regeneration pathways are shown in red.
Adenosine phosphoribosyltransferase catalyzes one of the primary salvage pathways. Here, free adenine reacts with PRPP to yield an adenine nucleotide:
\[\begin{equation} Adenine + PRPP \rightarrow AMP + PP_i \end{equation}\]
Free guanine and hypoxanthine (i.e., the deanimation product of adenine) are salvaged by hypoxanthine-guanine phosphoriboyltransferase.
12.5 Regulating De Novo Purine Biosynthesis
There are four key enzymes that are regulated:
- PRPP synthetase
- Amidophosphoribosyl transferase
- Adenylosuccinate synthetase
- IMP dehydrogenase
The first two enzymes regulate IMP synthesis; the last two regulate the production of AMP and GMP respectively.
12.5.1 Uric acid production from purine degradation
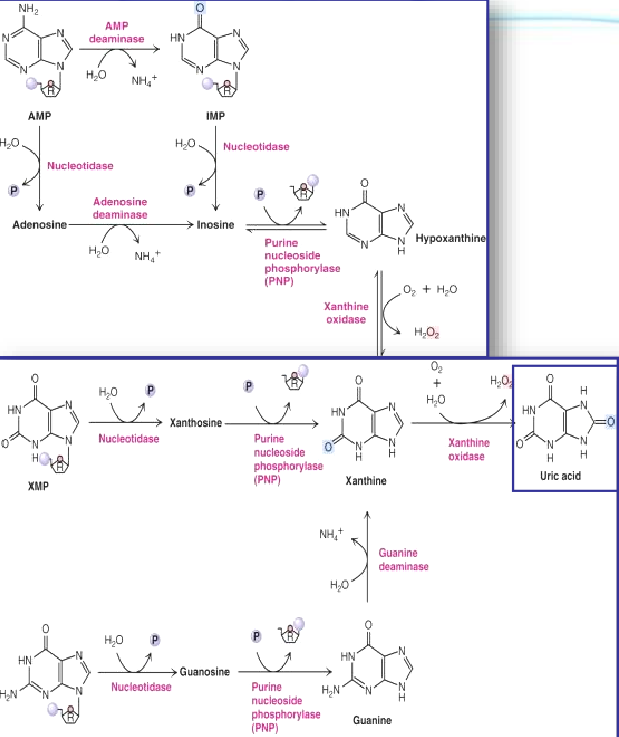
Figure 12.13: Uric Acid Production from Purine Degradation
Nucleosides are first degraded by purine nucleoside phosphorylase (i.e., PNP) to release the purine base and ribose-1-phosphate.
The PNP products are then merged into xanthine by guanine deaminase and xanthine oxidase. Xanthine is then oxidized into uric acid by the latter enzyme.
Xanthine oxidase is present in large amounts in the liver, the intestinal mucosa, and milk.
12.5.1.1 Gout from excessive uric acid accumulation
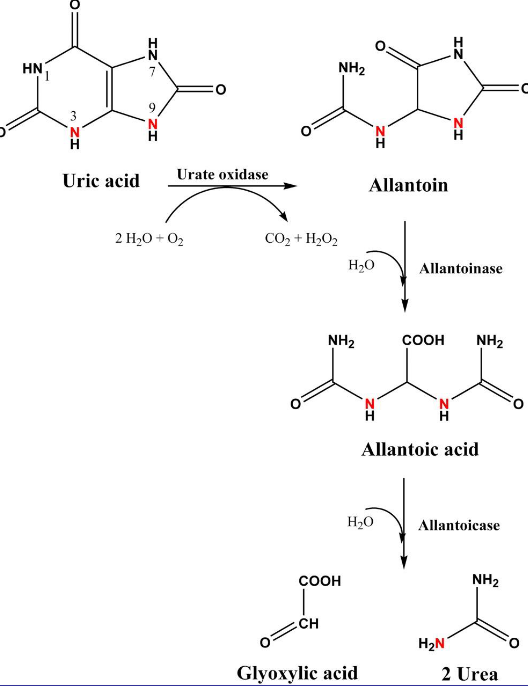
Figure 12.14: Urea Oxidase in Action
Uric acid can be converted into allatoin by urate oxidase. However, elevated levels of uric acid in the bloodstream can result in hyperuricemia.
Gout is a medical condition characterized by excessive amounts of uric acid in the body. Defects in PRPP synthetase and / or PRPP amidotransferase may render them insensitive to feedback inhibition by purine nucleotides. Consequently, too many nucleotides are produced and gout occurs.
Hypoxanthine-Guanine phosphoribosyltransferase (i.e., HGPRT) is a salvage pathway enzyme for purine metabolism.
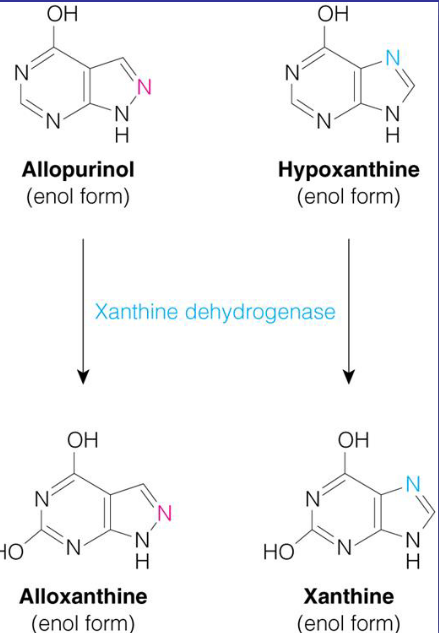
Figure 12.15: Xanthine Oxidase
Some gouts can also be treated with allopurinol - an inhibitor of xanthine oxidase
12.5.2 Lesch-Nyhan syndrome
In the absence or severe deficiency of HGPRT, activity leads to severe, gouty arthritis. The structural gene for HGPRT is located on the X-chromosome and is a recessive, sex-linked trait that only appears in males.
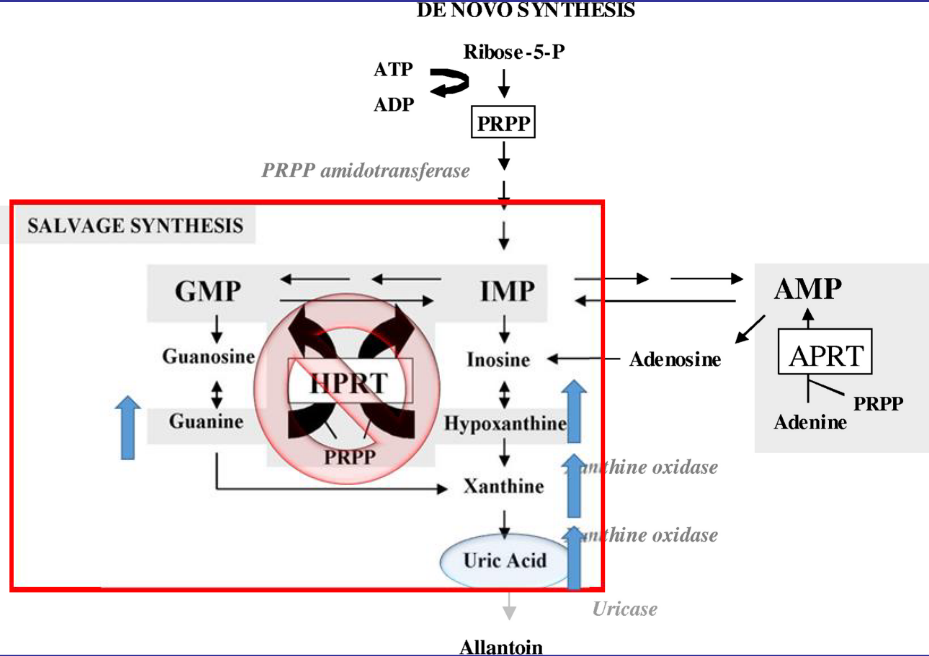
Figure 12.16: Mechanism Behind Lesch-Nyhan Syndrome
When HGPRT is absent, this leads to increased levels of uric acid in the blood. This is because de novo purine biosynthesis is dramatically increased.
12.6 Pyrimidines
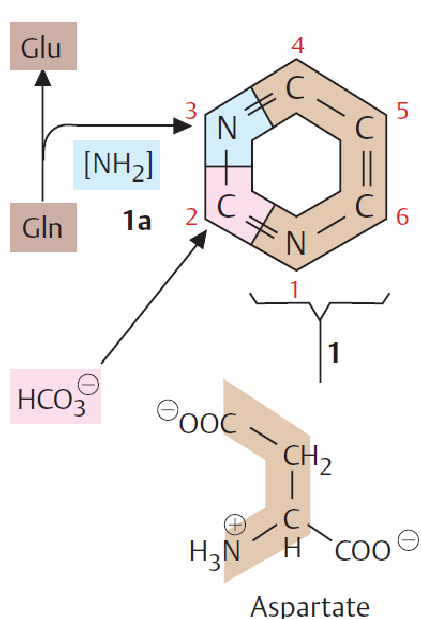
Figure 12.17: Pyrimidine Synthesis
Note that pyrimidines are not synthesized as nucleotide derivatives. The ring system is constructed prior to having a ribose-5-phosphate molecule attached to it.
12.6.1 Biosynthesis of pyrimidines
Such syntheses begin with the formation of carmoyl phosphate from carbamoyl phosphate synthetase-II (i.e., CPS-II)
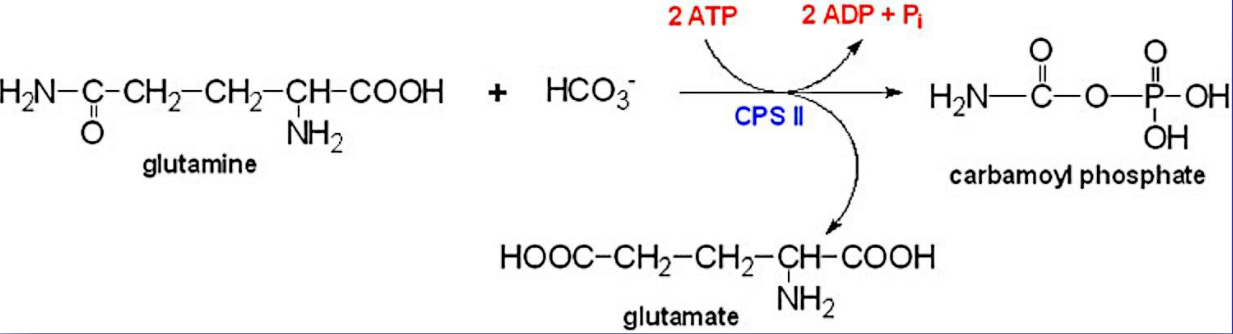
Figure 12.18: CPS-II Synthetase in Action
This molecule is formed from ATP, bicarbonate, and the amide nitrogen from glutamine.
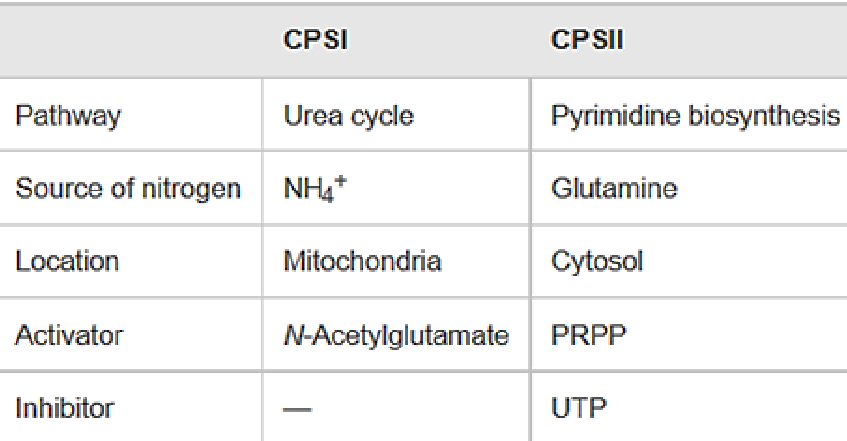
Figure 12.19: Differences Between CPS-I and CPS-II
Note the above table; CPS-II is unlike CPS-I from the Krebs-Henseleit urea cycle.
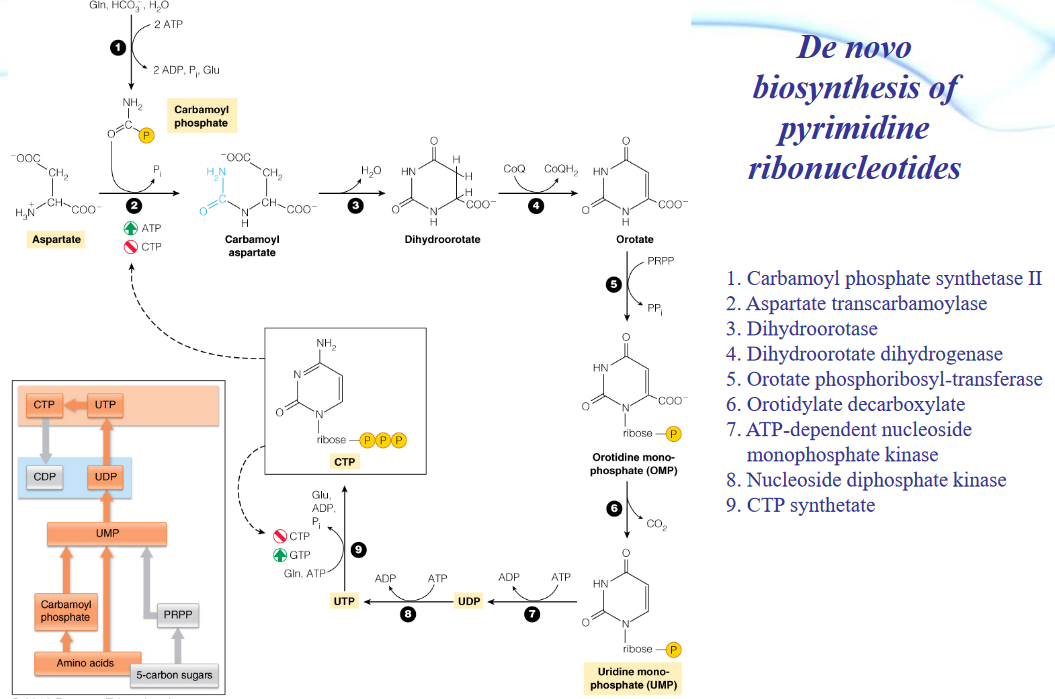
Figure 12.20: De Novo Synthesis of Pyrimidines
The biosynthesis of amino acids can also be summed up in the above graphic (taken from prof. Ardina’s slides).
12.6.2 Degradation of pyrimidines
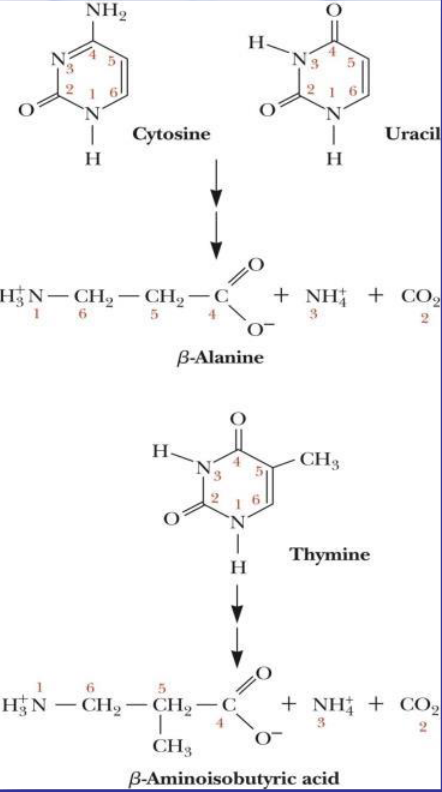
Figure 12.21: Degradation of Pyrimidines
Catabolizing cytosine and uracil yields \(\beta\)-alanine, ammonium, and CO2. Catabolizing thymine yields ammonium, CO2, and \(\beta\)-aminoisobutyric acid.
12.7 dNTP Biosynthesis
Ribonucleotides are precursors of deoxyribonucleotides. The pathway itself involves the reduction of ribose to deoxyribose (by replacing a C2 hydroxyl ion with a hydride ion) and the conversion of uracil to thymine.
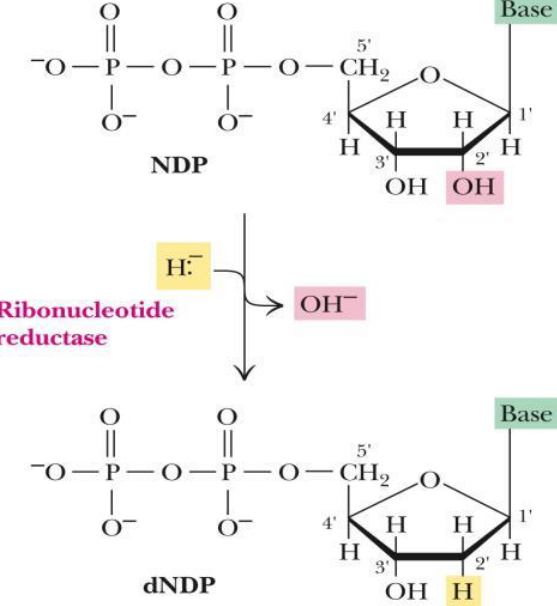
Figure 12.22: dNTP Biosynthesis
The enzyme ribonucleotide reductase (i.e., RNR) reduces all four common ribonucleotide substrates. Consequently, the enzyme is also oxidized.
12.7.1 RNR
There are three classes of RNR available:
- Class I uses a widespread, catalytic tyrosine residue.
- Class II uses a B12 coenzyme to generate radicals.
- Class III uses S-adenosylmethionine to generate a glycine radical.
RNR is an \(\alpha_2\beta_2\) heterodimer in E. coli, yeast, and humans.
12.7.2 Thioredoxin
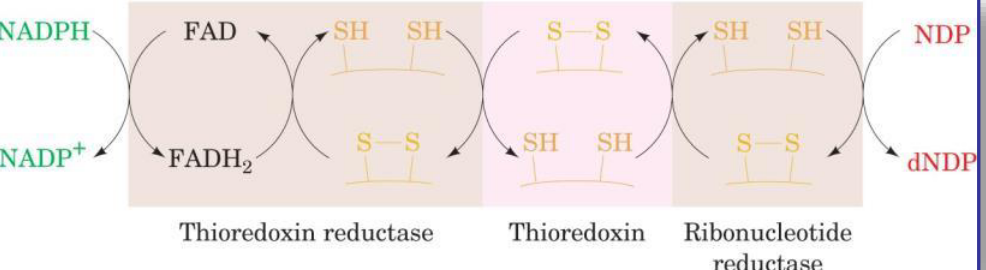
Figure 12.23: Thioredoxin
This molecule provides the reducing power for RNR.